
Les résultats décrits ci-dessous ont été tirés de la thèse soutenue en janvier 1996 (Cf. CV)
LE CRAWL
Introduction
Une des parts importantes de la formation en STAPS est liée à la connaissance et à l’initiation des pratiques techniques. En natation, comme dans de nombreux autres sports, la technique se réfère essentiellement à l’observation des plus grands champions. Afin de donner une assise scientifique à ces descriptions techniques, il est nécessaire de quantifier la gestuelle d’un grand nombre de nageurs de niveaux de performance différents, et en situation de compétition. Cette quantification permet alors de quantifier, d’infirmer et de compléter les descriptions techniques des ouvrages de référence.
L’étude des trajectoires des articulations du membre supérieur gauche va permettre de mettre en évidence les paramètres cinématiques et techniques déterminants du très haut-niveau de performance.
A partir de cette première étape de description de la gestuelle du nageur en crawl, les paramètres cinématiques ont été mis en relation avec la vitesse de nage, afin d’observer ceux qui avaient le(s) caractère(s) les plus discriminants de la performance.
Matériel et méthodes
Population :
Un échantillon de soixante nageurs masculins, de niveau national à international (Champion du Monde, 1994), réalisant un 100 m nage libre en compétition a été réalisé. La performance moyenne de ces nageurs sur le 100 mètres nage libre est de 1,89 m.s-1 (0,42 m.s-1) avec une étendue de 13,02 m.s-1.
Les valeurs moyennes de poids et de taille sont représentées dans le tableau suivant :
Tableau ?: Caractéristiques des sujets observés.
|
Paramètres |
Moyenne |
Écart type |
Etendue |
|
Poids (kg) |
78,94 |
15,02 |
31,5 |
|
Taille (m) |
1,847 |
0,11 |
0,25 |
|
Age (année) |
21,4 |
6,29 |
11 |
|
Perf.(100 m) (s) |
52,79 |
11,73 |
13,02 |
L'homogénéité de notre population du point de vue de la taille nous permet de neutraliser ce facteur dans l’identification des paramètres de la trajectoire relatifs au haut-niveau de performance.
Epreuve
Les enregistrements vidéo ont été réalisés durant les Championnats de France d'hiver 1993 et 1994, lors de la Coupe du Monde à Paris (1994), ainsi que lors des Championnats du Monde à Rome (1994), lors des séries et des finales du 100 mètres Nage Libre (NL) messieurs.
Le 100 m NL a été choisi d’une part parce que cette course est celle qui comprend le plus de nageurs lors des compétitions. La recherche des paramètres discriminants du niveau de performance en crawl rendait indispensable le choix d’un très grand nombre de nageurs de niveaux hétérogène. D’autre part, la partie filmée lors des épreuves étant située dans les derniers 20 m du premier cinquante mètres de nage de la course, les nageurs avaient effectué environ 40 % de leur course. Il semble alors que la fatigue ne puisse être un facteur susceptible d'entraîner des variations dans nos résultats, surtout lors de cette compétition où les sujets sont supposés être en forme optimale (Hay and Guimaraes, 1983; Pai et al., 1984; Schleihauf et al., 1988).Acquisition des données
Les outils utilisés devaient nous permettre d’acquérir des prises de vue sous-marines en cours d’épreuve. Les impératifs du logiciel d’analyse cinématique développé par R.E. Schleihauf (1994) nous imposent d’avoir deux caméras synchrones formant un angle de 90° entre elles.
Deux camescopes (Hi8 modèle EVO 150 TR, format NTSC, avec time code intégré; annexe 1) de 30 Hz chacun ont été utilisés. Cette fréquence de 30 Hz est largement suffisante pour étudier un cycle aquatique des nageurs, celui-ci ayant une durée moyenne d'environ 1 seconde.
Compte-tenu de ces conditions d'acquisition, les réglages des camescopes étaient manuels afin d'éliminer toute adaptation automatique durant les prises de vues. En particulier, nous avons neutralisé le focus automatique afin d’éviter un éventuel changement d'échelle de référence à tout instant. La vitesse d'obturation était de 1/250, compte tenu de la faible luminosité en vues sous-marines dans une piscine et de la vitesse relativement lente du mouvement des nageurs. Aucun effet de zoom n'a été utilisé, afin de garder un champ de vision en moyenne de 6 mètres. Ce champ nous a paru suffisant compte-tenu de l'amplitude d'un cycle de bras aquatique (environ 2 mètres) (Craig et Pendergast, 1979; Craig et al., 1985; Maglischo, 1986 et al.; Chollet, 1991; Chollet et Pelayo, 1995) et de la taille moyenne des sujets (1,809 m). Ce réglage permet d'obtenir environ deux cycle de bras pour chaque nageur filmé.
Pour pallier aux problèmes d'étanchéité, chacun des deux camescopes a été installé dans un caisson étanche (type Sony SPK-TRA). Dans chacun des caissons, nous avons également placé des petites diodes lumineuses, nous permettant de synchroniser les enregistrements entre les deux camescopes.
Du fait du placement des camescopes sous l'eau dans leur caisson et de la non possibilité de pouvoir les sortir en cours de compétition, un système d'alimentation externe d'une autonomie de plus de six heures a été utilisé (batteries 12 V, à une puissance de 10 Ah). Un câble étanche étant relié aux caissons permettait ainsi l’alimentation des camescopes de l’extérieur du bassin, mais contenait également d’autres câbles tels que celui de la télécommande de déclenchement vidéo (déclenchement simultané des deux camescopes), celle de démarrage des diodes de synchronisation, ainsi que celui permettant d'obtenir un retour vidéo durant toute la durée de l'enregistrement (vérification à chaque instant des prises de vues grâce à un petit écran Sony (LCD Color Monitor, XV-M30).
Le placement de nos caméras devait tenir compte des contraintes réglementaires imposées par la Fédération Internationale de Natation Amateur (F.I.N.A.) et des impératifs liés au logiciel "Kinematic Analysis" de R.E. Schleihauf.
Le traitement des images, effectué à partir du logiciel développé par R.E. Schleihauf (1994), nécessite d'avoir un angle de 90° entre les champs des deux camescopes. De part le fait de filmer lors de compétitions, il était impossible de placer les camescopes de face et de profil par rapport au déplacement du nageur. Ils ont ainsi été placés avec un angle de 45° par rapport aux axes de déplacement des nageurs (FIGURE), dans les deux angles de la piscine, côté virage.
Les ensembles "camescope et caisson" ont été vissés à 45° sur une équerre horizontale en Plexiglas, elle-même vissée sur une plaque verticale de Plexiglas plaquée contre le mur grâce à des lests.
Le positionnement en angle nous a donné une distance entre l'objectif des camescopes et le champ étudié de 8 m. De par les problèmes de convergence de champs des deux camescopes, seuls les nageurs évoluant dans les deux couloirs centraux ont pu être analysés. Chacun des deux ensembles (camescopes+caissons) a été positionné à 0,60 m sous la surface de l'eau, distance nécessaire compte-tenu de la profondeur maximale atteinte par la main lors d'un cycle aquatique (FIGURE).
Les batteries, les moniteurs vidéo de contrôle, ainsi que les télécommandes se trouvaient au maximum à 10 m du bord du bassin, sur une surface sèche, et disposés de telle façon à ce qu’un seul opérateur puisse contrôler l’ensemble des acquisitions.
Figure ?: Représentation schématique du placement des caissons lors des compétitions
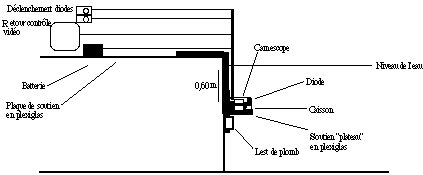
Figure ?: Placement de l'ensemble "caisson et camescope" par rapport au bord du bassin (vue de profil).
Traitement des données
A partir des prises de vue sous-marines, une digitalisation image par image de certaines articulations du corps du nageur a été effectuée selon le logiciel développé par Schleihauf R.E. (Nouvelle version, 1994). Le processus de digitalisation comprend plusieurs étapes :
- Visualisation des images:
Dans un premier temps, les cassettes Hi8 enregistrées sur les camescopes sont visualisées à partir d’un magnétoscope Sony (EVO-9650) sur un moniteur Sony, d'une définition de 230 lignes TV permettant de visionner les images. Le magnétoscope Sony est un matériel de type professionnel possédant une molette de recherche, permettant de faire défiler l'enregistrement image par image. Un time code intégré au trentième de seconde permet de repérer facilement les séquences du film à exploiter. Le cycle de bras à digitaliser est alors sélectionné parmi les vues des deux caméras. Pour le crawl, un cycle de bras a été décrit entre deux entrées successives de la main gauche dans l'eau. Nous avons délibérément choisi de travailler sur un cycle pendant lequel le sujet n’inspire pas, les mouvements de la tête et du tronc lors de l’inspiration pouvant avoir une influence sur la trajectoire du bras
- Capture des images:
Dans un deuxième temps, une saisie informatique est effectuée pour chacune des images correspondant au cycle à étudier. La numérisation des images vidéo est réalisée grâce à une carte d’acquisition TARGA. Cette carte permet également un retour des images numérisées sur écran vidéo. Les images numérisées sont stockées dans le disque dur d’un ordinateur PC d'une capacité de 200 Mb.
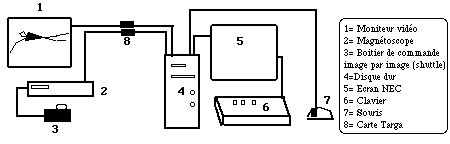
Figure 9: Les différents outils permettant le traitement des données
- Digitalisation: La digitalisation est ensuite effectuée à partir des images numérisées. En crawl, en accord avec la méthode de Schleihauf, nous avons créé un modèle comprenant 17 points de référence sur le corps du nageur. Ceux ci sont pointés, pour chacune des images des deux caméras, à l’aide d’une souris raccordée à l’ordinateur :
- Les bout des majeurs droit et gauche - les hanches droite et gauche
- les poignets droit et gauche - les genoux droit et gauche
- les coudes droit et gauche - les chevilles droite et gauche
- les épaules droite et gauche - les orteils droit et gauche
- la base du cou
Les coordonnées en 2 dimensions (2D) de chacun des points sont visualisées sur un moniteur RGB analogue graphique (Sony Trinitron, 512X400 pixels, 32000 nuances de couleur) et stockées dans le disque dur de l'ordinateur. Pour établir les coordonnées des points digitalisés en référence à un repère fixe externe au nageur, un point de référence (0,0) est également digitalisé sur chacune des images. Pour permettre de connaître l’échelle nous permettant de calculer l’amplitude des déplacements du nageur, une image de référence est également utilisée à la fin de chaque séquence de digitalisation. Cette image de référence correspond à la digitalisation d’une mire de 1,30 m de longueur.
Après la digitalisation, les coordonnées des points sont obtenues pour chaque plan des deux caméras (nous obtenons donc deux fois deux dimensions). Les impératifs liés à la situation de compétition, (placement des caméras en angle) nous donnent des coordonnées pour chacune des 3 dimensions correspondant aux plans des caméras(45° par rapport au déplacement du nageur). Une rotation de 45° a donc été effectuée sur ces coordonnées par une fonction de vissage pour obtenir les trajectoires des points digitalisés pour chacune des dimensions en référence à un repère externe :
- la direction antéro-postérieure donne les variations "avant-arrière" des coordonnées. Lorsque le sujet avance sur cet axe par rapport au point de référence externe, les coordonnées vont augmenter
- la direction transversale donne les variations "interne-externe" des coordonnées. Lorsque le sujet se déplace vers l'intérieur par rapport au point de référence, les coordonnées augmentent ;
- la direction verticale donne les variations "haut-bas" des coordonnées. Lorsque le sujet se déplace vers le haut par rapport au point de référence, les coordonnées vont augmenter.
- Lissage:
Comme toute donnée biomécanique (Dainty & Norman, 1987) les trajectoires obtenues doivent être lissées pour minimiser les erreurs liées la digitalisation. Les données recueillies sont donc lissées à partir d'une fonction polynomiale de degré trois. Le choix de la fonction polynomiale comme outil de lissage a été réalisé par R.E. Schleihauf.
Exportation des données :
Nos logiciels d’analyse de données (univariée, bivariée, multivariée dont ACP) étant sur Macintosh, nous avons du exporter nos données pour effectuer leur traitement statistique.
La durée du cycle de bras n'est pas toujours la même selon les nageurs. Pour comparer les phases et les trajectoires des différents nageurs, nous avons effectué une normalisation temporelle de leur cycle. Pour chaque nageur, la durée du cycle aquatique représente alors 100%t du temps. Les coordonnées de chacune des articulations sont alors obtenues pour chaque 5% du temps du cycle aquatique.
Par ailleurs, le cycle de bras étudié pour chaque nageur ne commence pas strictement au même endroit dans le champ des caméras. De ce fait, par exemple, la première coordonnée (entrée de la main dans l’eau), sur l’axe antéro-postérieur, en référence à un repère externe sera différente d’un nageur à l’autre. Pour pouvoir comparer nos nageurs nous avons donc du réaliser une normalisation spatiale. Nous avons alors effectué un changement d’origine pour chacune des trajectoires de chacune des articulations digitalisées. Chaque articulation a sa propre origine (0,0,0) qui correspond à sa position au moment de l'entrée dans l'eau.
A partir de cette normalisation, nous avons pu effectuer les trajectoires moyennes représentant nos 60 nageurs pour chacune des articulations étudiées.
Paramètres étudiés
Dans un premier temps, nous avons calculé la vitesse de déplacement de la hanche gauche (Vh) sur l’axe antéro-postérieur.
Vh (m/s)= Dh (m)/Tc (s)
où Dh = (coordonnée de la hanche à la sortie de la main de l’eau-coordonnée de la hanche à l’entrée de la main dans l’eau),
Tc = temps de cycle aquatique (temps entre l’entrée et la sortie de la main de l’eau).
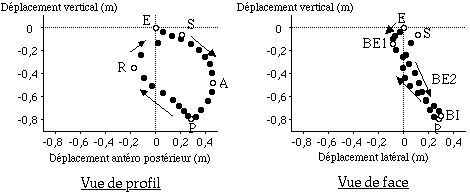
Dans un deuxième temps, dans le but de décrire la technique de nage des nageurs, nous avons observé la trajectoire moyenne de leur poignet sur un cycle aquatique complet, sur les trois axes : antéro-postérieur, transversal et vertical. L’amplitude et la durée de ces trajectoire ont été relevées. L’amplitude correspond alors au déplacement du poignet entre son entrée (E) et sa sortie (S) de l’eau, et la durée est celle du cycle aquatique.
- Enfin, nous avons observé, toujours dans l’objectif de décrire plus finement la technique de nage, les différentes phases du cycle aquatique du poignet par rapport à un point de référence externe au nageur (TABLEAU et FIGURE).
|
PHASES |
AXES |
||
|
Antéro-postérieur |
Transversal |
Vertical |
|
|
Avancée (A) |
CoordS-CoordA |
||
|
Recul (R) |
CoordA-CoordR |
||
|
1er balayage (BE1) |
(Coord +Ext.)-CoordE |
||
|
Balayage interne (BI) |
(Coord+Int.)-(Coord +Ext.) |
||
|
Balayage externe(BE2) |
CoordS-(Coord.+Int.) |
||
|
Prof. au point A (PA) |
Coord. au point A |
||
|
Profondeur max. (Pmax) |
Coord.mini |
Les amplitudes et les durées de ces différentes phases ont été calculées en moyenne pour l’ensemble des sujets de l’étude.
Traitement statistique
Les moyennes et les écarts types ont été calculés pour chacune des coordonnées des trajectoires de la hanche et du poignet, ainsi que pour les différentes amplitudes et durées du cycle aquatique et de ses phases.
Résultats
1) Vitesses de nage et vitesses de cycle
En natation, et en crawl plus précisément, le but recherché est de parcourir une distance donnée dans le minimum de temps. "Dans son principe, il constitue l'essence supérieure de la natation sportive, puisqu'il incite les nageurs à toujours réaliser de meilleurs temps .../...Le but de la course, c'est d'aller le plus vite possible..." (Oppenheim, 1977). La vitesse de nage est un indicateur du niveau de performance des sujets. Dans notre étude, la vitesse de nage moyenne sur le 100 mètres est de 1,89 m.s-1 (0,42 m.s-1). Notre population est dispersée pour ce paramètre. Le crawl étant une nage cyclique, la vitesse de nage est la résultante de la vitesse de déplacement du corps au cours de chaque cycle aquatique de bras.
Afin d’évaluer la représentativité du cycle aquatique étudié dans notre étude par rapport à la globalité du 100 mètres, nous avons mis en relation la vitesse moyenne sur 100m avec la vitesse du corps au cours du cycle de bras étudié. La plupart des auteurs s'accordent pour assimiler la vitesse horizontale du corps au niveau de performance du nageur (Counsilman, 1968; Counsilman, 1977; Barthels, 1977; Maglischo et al., 1986; Maglischo, 1987; Chollet, 1991). Certains auteurs ont montré que la vitesse de déplacement de la hanche au cours d'un cycle était équivalente à la vitesse de déplacement du centre de gravité et donc du corps (Counsilman, 1968; Craig et Pendergast, 1979; Hay and Guimaraes, 1983). Au cours de notre étude, nous assimilerons donc la vitesse du corps à la vitesse de la hanche.
La vitesse moyenne de la hanche sur l'axe antéro-postérieur lors du cycle aquatique est de 1,85 m.s-1 (0,22 m.s-1). La population est relativement dispersée pour ce paramètre avec des valeurs comprises entre 1,30 m.s-1 et 2,29 m.s-1 (FIGURE).
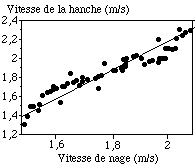
Figure ?: Relations entre la vitesse de nage et la vitesse de la hanche sur le cycle aquatique étudié
Une relation linéaire significative (r = 0,81 p = 0,001) est montrée entre la vitesse de la hanche sur l'axe antéro-postérieur et la vitesse de nage (calculée à partir du temps réalisé sur le 100 m). Les nageurs les plus rapides sur le 100 m présentent les vitesses horizontales de la hanche les plus élevées au cours du cycle. Le cycle étudié est donc bien représentatif de la course du point de vue de la vitesse.
La vitesse étant le rapport de l'amplitude de déplacement et la durée de cycle, nous avons étudié dans un premier temps les relations entre le niveau de performance et les paramètres temporels. Les meilleurs nageurs ont-ils un temps de cycle plus court? Présentent-ils des différences temporelles au niveau des phases aquatique et de retour aérien?
2) Etude temporelle du cycle de bras
les durées d’un cycle total compris entre deux entrées successives de la main gauche dans l’eau, ainsi que les durées des phases aquatique et aérienne de la même main ont été étudiées.
2.a) Durée totale du cycle
La durée moyenne d’un cycle, c’est à dire entre deux entrées successives de la main gauche dans l’eau est de 1,47 s (0,16 s). Les sujets sont répartis de manière homogène autour de la moyenne, ce qui signifie qu’ils n’enregistrent pas de durées très différentes de leur cycle de bras. Ces données sont légèrement supérieures à celles obtenues par des études antérieures telles que Renner (1977) et Craig et al. (1985).
2.b) Durées des phases aquatique et aérienne
La phase aérienne, définie entre la sortie de la main gauche de l’eau et son entrée dans l’eau consécutive est en moyenne de 0,65 s (0,06 s) pour notre population. Elle est significativement (p = 0,009) plus courte que la phase aquatique, définie entre l’entrée de la main gauche dans l’eau et sa sortie consécutive (0,81 s (0,08 s)). Précédemment, certains auteurs avaient montré des valeurs beaucoup plus courtes de cette phase aquatique (Miyashita 1975 observait une valeur moyenne de 0,58 s par exemple). Cette différence pourrait provenir de l’antériorité de l’étude, mais également du type de test réalisé (25 yard).
Rapportées au temps total du cycle de bras, les valeurs moyennes de la phase aquatique sont de 55,4 % (5,01 %) contre 44,5 % (3,98 %) pour la phase aérienne.
Comme pour la durée totale du cycle, les sujets que nous avons observé restent très homogènes autour de la moyenne, quelle soit la partie du cycle étudiée. La stabilité temporelle des différentes phases d’un mouvement pourrait être à ce niveau de performance une constante du mouvement. En effet, dans le domaine de la performance motrice, des études avaient auparavant montré beaucoup plus d’invariances temporelles que spatiales (Schmidt et al., 1979 ; Shapiro et al., 1981). L’acquisition d’une habilité motrice suppose qu’un nombre le plus réduit possible de paramètres doivent rester sous le contrôle du système nerveux (Bernstein, 1967 ; Schoebel et Hay, 1990). Ces invariants permettraient de réduire le coût du système nerveux. De plus, les structures temporelles seraient plus résistantes que les structures spatiales, et constitueraient alors une sorte d’invariants de l’habilité motrice. En d’autres termes, le temps serait " la composante la plus invariante " qui sert dans une certaine mesure de cadre stable autour duquel s’effectuent d’autres adaptations, notamment des adaptations spatiales.
La vitesse étant le rapport entre l’amplitude et le temps de cycle, et les nageurs n’étant pas différents du point de vues de leurs différentes durées de cycle, l’influence des facteurs spatiaux est alors étudiées, puis mis en rapport avec la vitesse de nage.
3) Description de la technique de nage en crawl
Pour l’études des trajectoires, seule la partie aquatique du cycle du bras gauche a été étudiée. Les trajectoires du poignet, du coude, de l’épaule et de la hanche sont présentées FIGURE ?.
Les trajectoires ne sont pas linéaires dans chacune des trois dimensions : antéro-postérieure, transversale et verticale. Cela signifie que les articulations enregistrent des mouvements avant-arrière, droite-gauche et haut-bas entre les deux points d’entrée et de sortie de l’eau, définissant le cycle aquatique. Sur l’axe antéro-postérieur, lorsque les coordonnées augmentent, l’articulation considérée se dirige vers l’avant et réciproquement. Sur l’axe transversal, lorsque les coordonnées augmentent, l’articulation se dirige vers l’intérieur par rapport à l’alignement du corps. Et enfin une diminution des coordonnées sur l’axe vertical définit un mouvement vers le bas de l’articulation.
Les amplitudes de déplacement du cycle aquatique ainsi que celles des paramètres définis dans la partie protocole ont été observées.
3.a) Trajectoires moyennes
La figure suivant permet d’observer les trajectoires moyennes du poignet recueillies en moyenne chez nos 60 sujets de profil (figure de gauche) et de face (figure de droite).
Dès son entrée dans l’eau, le poignet se dirige simultanément vers l’avant, vers le bas et légèrement vers l’extérieur. Cette première phase est appelée phase de prise d’appui par la plupart des auteurs. Nous observons, qu’à la différence de la trajectoire de la main décrite par Maglischo (1984) par exemple (FIGURE), le poignet se dirige simultanément vers l’avant et vers le bas dans notre étude, et non successivement vers l’avant puis vers le bas.

Figure : Trajectoire sagittale de la main décrite par Maglischo (1984)
Après cette phase de prise d’appui, le poignet se dirige vers le bas et vers l’intérieur. Cette phase définit la phase de traction expliquée par d’autres auteurs. Nous remarquerons que cette phase se termine après l’atteinte du point " P " le plus profond de la trajectoire, alors que l’articulation n’a pas encore tout à fait atteint le point le plus interne (BI).
Enfin, après avoir terminé son balayage vers l’intérieur (BI), le poignet se dirige vers l’arrière puis vers l’arrière et le haut. Cette phase définie la phase de recul de l’articulation. Comme en début de cycle aquatique, nous remarquerons une différence au niveau de la trajectoire décrite antérieurement par Maglischo et celle observée dans notre étude. Alors que Maglischo distinguait un mouvement de type " sinusoïdal " de la main, nous décrivons une trajectoire beaucoup plus rectiligne, directement vers l’arrière et le haut lors de cette dernière phase de poussée.
De grosses différences apparaissent entre les nageurs au niveau notamment de certaines phases du cycle du poignet telles que la première partie du mouvement et la phase de recul. Les amplitudes de déplacement ont donc été observées pour chacune des phases du cycle aquatique.
3.b) Les différentes phases du cycle aquatique
TABLEAU : Moyennes et écarts types des amplitudes de déplacement et durées relatives (en italique) du poignet et du coude sur chacun des points caractéristiques de leur cycle aquatique. Moyennes et écarts types des amplitudes de déplacement antéro-postérieures de l’épaule et de la hanche lors de l’atteinte par le poignet des points caractéristiques de sa trajectoire aquatique.
|
Ampli. lors des points caractér. |
Ampli. ant.post. lors des points caract. |
|||
|
Poignet |
Coude |
Epaule |
Hanche |
|
|
A |
0,5m (0,27m) 43,4% (10,2%) |
0,61m (0,3m) 58,3% (20,8%) |
0,61m |
0,55m |
|
P |
-0,7m (0,1m) 64% (8,5%) |
-0,4m (0,07m) 62,5% (8,07%) |
0,90m |
0,80m |
|
R |
-0,65m (0,18m) 51,1% (9,9%) |
-0,19m (0,1m) 27,7% (2,3%) |
1,30m |
1,30m |
|
BE1 |
-0,08m (0,06m) 50% (8,01%) |
0,21m (0,09m) |
0,19m |
0,19m |
|
BI |
0,16m (0,05m) 20,8% (4,3%) |
66,7% (9,1%) |
0,98m |
0,86m |
|
BE2 |
-0,23m (0,11m) 29,2% (5,5%) |
0,26m (0,11m) 43,3% (7,5%) |
1,38m (0,25m) |
1,48m (0,17m) |
-Sur l’axe antéro-postérieur :
La hanche et l’épaule enregistrent des déplacements relativement réguliers vers l’avant durant le cycle aquatique du bras gauche (FIGURE).
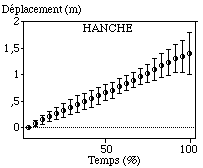

Figure: Trajectoires antéro-postérieures moyennes de la hanche et de l'épaule sur le cycle aquatique
Leurs amplitudes respectives sont 1,48 m (0,17 m) et 1,38 m (0,25 m). La différence de déplacement entre ces deux articulations provient du mouvement propre de l’épaule lors de l’entrée du bras dans l’eau (extension vers l’avant), et de sa sortie de l’eau (du fait de la poussée de la main en arrière).
Le poignet et le coude par contre se déplacent vers l’avant puis vers l’arrière lors de leurs cycles aquatique (FIGURE).
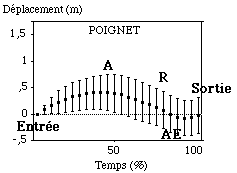
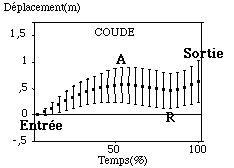
Figure : Trajectoires antéro-postérieures moyennes du poignet et du coude sur le cycle aquatique
Bien que le poignet sorte à peu près à l’endroit où il était entré dans l’eau (amplitude de déplacement totale lors du cycle aquatique de -0,01 m (0,33 m), il enregistre donc des variations très importantes vers l’avant et vers l’arrière durant son cycle. Le mouvement initial du poignet après l’entrée de la main dans l’eau est décrit par un déplacement important vers l’avant (" A " = 0,5 m (0,27 m)) durant près de 44 % (10,2 %) du cycle aquatique. Les sujets sont très dispersés autour de la moyenne quant à ce déplacement. Cela signifie qu’ils n’enregistrent pas tous la même forme et la même amplitude de mouvement. Après ce mouvement, le poignet se dirige vers l’arrière par rapport à cet axe antéro-postérieur. Cette phase appelée pour notre étude " phase de recul " a une amplitude de -0,73 m (0,1 m). L’articulation va donc ainsi passer en arrière de la coordonnée d’entrée dans l’eau, par rapport à un repère externe au nageur. Cette phase de recul est plus longue que la phase initiale d’avancée vers l’avant (64 % (8,5 %) du cycle aquatique).
Globalement le poignet va sortir en avant du point où il est entré dans l’eau (0,74 m (0,2 m)). Par contre, durant son cycle aquatique, le coude enregistre le même type de mouvement que le poignet mais avec des amplitudes différentes de l’ordre de 0,6 m (0,3 m) pour la phase d’avancée initiale et de -0,4 m (0,1 m) pour la phase de recul. Cette articulation " avance " donc de manière plus importante en début de cycle aquatique, et observe une phase de recul moins importante, ne passant jamais en arrière de sa coordonnée d’entrée dans l’eau. Contrairement à ce qui fut décrit dans les livres à caractère de description technique, la " fuite " du coude vers l’arrière est donc moins importante du point de vue de son amplitude déplacement que celle du poignet (Maglischo, 1984). Les durées de ces phases d’avancée et de recul sont sensiblement les mêmes pour le coude que pour le poignet, légèrement supérieure pour le coude lors de la phase d’avancée en début de cycle aquatique. Le poignet ayant alors un mouvement propre autour du coude qui servirait alors d’axe de rotation ?
Lors de la phase d’avancée (définie par la trajectoire décrite par le poignet entre son entrée dans l’eau et le point " A "), la hanche avance de 0,55 m (0,31 m). La grande dispersion des sujets au niveau de cette amplitude du corps en début de cycle aquatique montre que certains nageurs ont la possibilité d’obtenir par le(s) mouvement(s) du membre supérieur un plus grand déplacement du corps en ce tout début de cycle aquatique. A la fin de la phase de recul, le corps a avancé de 1,30 m (0,41 m) au total, donc de 0,75 m entre le point " A " et le point " R ". Il semblerait que ce n’est pas uniquement le mouvement vers l’arrière du poignet qui permette cette avancée, mais également ses mouvements transversaux et verticaux, ainsi certainement les mouvements simultanés du bras opposé et des jambes.
-Sur l’axe transversal :
La hanche et l’épaule enregistrent de très faibles déplacements transversaux lors des cycles aquatiques du bras gauche pour notre population (de l’ordre de 0,10 m en moyenne) (FIGURE).

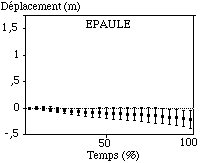
Figure: Trajectoires transversales moyennes de la hanche et de l'épaule sur le cycle aquatique
Le poignet et le coude par contre enregistrent des déplacements vers l’extérieur et vers l’intérieur par rapport au corps lors de leurs cycles aquatiques. Contrairement à l’axe antéro-postérieur, ces mouvements transversaux sont différents pour les deux articulations (FIGURE).
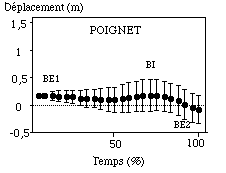

(*) la valeur (0) sur l’axe des abscisses correspond à la position de l’épaule lors de l’entrée du poignet dans l’eau
Figure : Trajectoires transversales moyennes du poignet et du coude lors du cycle aquatique
Le poignet commence tout d’abord par une phase de balayage vers l’extérieur (BE1, autrement appelée phase de prise d’appui) d’amplitude relativement faible (-0,08 m (0,06 m)), mais de durée très importante (50 % (8,01 %) du cycle aquatique). Ensuite, l’articulation revient vers l’intérieur, c’est à dire sous l’axe du corps, ce qui définit le balayage interne (BI) d’amplitude et de durée respectivement égales à 0,16 m (0,05 m) et 20,8 % (4,3 %) ; pour terminer le cycle aquatique par un second balayage vers l’extérieur (BE2) d’amplitude de 0,2 m (0,04 m). Nous notons que les amplitudes de déplacements du poignet sur cet axe transversal sont très nettement inférieures à celles observées sur l’axe antéro-postérieur. Le poignet se dirige donc principalement vers l’avant et l’arrière, et beaucoup moins de l’extérieur vers l’intérieur et réciproquement.
Le coude n’enregistre pas réellement de balayage externe en début de cycle aquatique. Comme sur l’axe antéro-postérieur, il servirait plus principalement d’axe de rotation relativement fixe autour duquel le poignet effectue sa phase de prise d’appui vers l’extérieur. Son mouvement principal s’oriente majoritairement vers l’intérieur (0,21 m (0,09 m)) durant 67 % (9,1 %) du cycle aquatique ; pour ensuite terminer par un balayage vers l’extérieur afin de se " dégager " pour sortir de l’eau.
Les amplitudes de déplacements antéro postérieur du corps ont été observées durant ces phases transversale du poignet. Lors du premier balayage externe, le corps avance 0,19 m, puis à la fin du balayage vers l’intérieur l’amplitude de déplacement est de 0,86 m (c’est à dire une avancée brute entre BE1 et BI de 0,67 m (0,21 m) en moyenne). Comme sur l’axe antéro-postérieur, les sujets sont très différents quant à l’avancée globale de leur corps durant les différents balayages de leur poignet, notamment lors de la phase de balayage interne.
-Sur l’axe vertical :
Comme sur l’axe transversal, la hanche et l’épaule n’enregistrent pas de mouvements très importants vers le haut et vers le bas (de l’ordre de 0,18 m). Ces seuls mouvements, notamment au niveau de l’épaule n’ont uniquement pour objectif de dégager le coude et le poignet, afin d’augmenter éventuellement leurs amplitudes de déplacement (FIGURE).

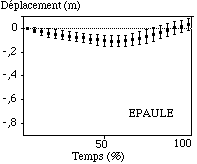
Figure: Trajectoires moyennes de la hanche et de l'épaule sur l'axe vertical
Le poignet et le coude enregistrent quant à eux de gros déplacements verticaux, définis sous la forme de deux phases distinctes : la première entre l’entrée dans l’eau et le point le plus profond, et la seconde par la remontée vers le haut de l’articulation (entre le point " P " et la sortie de l’eau)(FIGURE).
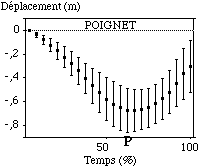
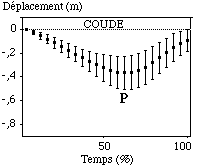
Figure : Trajectoires verticales moyennes du poignet et du coude sur le cycle aquatique
Après l’entrée dans l’eau, le poignet se dirige vers le bas. Lors de l’étude générale des trajectoire, nous avions insister sur le fait qu’en début de cycle aquatique, le poignet se dirigeait simultanément vers l’avant et vers le bas. L’amplitude de la profondeur de l’articulation lors du point " A " confirme ces propos (0,48 m (0,24 m)). Lors du point d’avancée maximale vers l’avant (" A "), le poignet s’est donc déjà enfoncé d’environ 0,50 m, décrivant bien un mouvement concomitant vers l’avant et vers le bas. Les sujets sont très hétérogènes quant à cette valeur. Leur capacité à déplacer le poignet très rapidement vers l’avant et vers le bas en début de cycle est donc très différente. La profondeur maximale atteinte par le poignet est de -0,73 m (0,1 m) à 64 % (8,5 %) du cycle aquatique. Plus de la moitié du cycle aquatique est donc dépensée à " descendre " l’articulation vers le bas. Nous notons une nouvelle fois une très grosse dispersion des sujets quant à ce paramètre.
Le coude enregistre environ le même mouvement que le poignet sur cet axe vertical, atteignant une profondeur maximale de -0,4 m (0,07 m) à peu près en même temps que le poignet (à 63 % (8,07 %) du cycle aquatique).
Durant la phase initiale d’enfoncement des articulations, le corps avance de 0,80 m (0,38 m) en moyenne chez nos sujets, alors que lors de la seconde phase, lorsque les articulations du poignet et du coude remontent à la surface de l’eau, le corps ne va avancer que de 0,68 m. La première phase du cycle aquatique va donc permettre en majorité chez les nageurs observés un plus grand déplacement vers l’avant de l’ensemble du corps. Les sujets sont une nouvelle fois très dispersés autour de la moyenne, ce qui indique une amplitude très différente de leur déplacement en ce début de cycle aquatique.
4) Relation entre paramètres cinématiques et niveau de performance
Lors du chapitre précédent : " description de la technique de nage en crawl ", nous avons pu observer que les nageurs se révélaient hétérogènes quant aux valeurs de certains de leurs amplitudes de déplacement. Ce chapitre va permettre de mettre en relation ces différents paramètres de déplacement avec la vitesse de nage, afin d’observer si cette hétérogénéité des sujets pouvait éventuellement être expliquée par le niveau de performance de nos nageurs.
Le premier résultat que nous pouvons mettre en évidence est le rapport évident existant entre la vitesse de nage de nos sujets et l’amplitude de déplacement de leur hanche sur l’axe antéro-postérieur, c’est à dire l’avancée globale de leur corps au cours du cycle aquatique. Un coefficient de corrélation linéaire de r = 0,82 p = 0,0002 est obtenu lors du croisement entre la vitesse de nage et l’amplitude de déplacement de la hanche lors du cycle aquatique sur l’axe antéro-postérieur (FIGURE). Cela signifie que les meilleurs nageurs enregistrent également la plus grande avancée de leur corps vers l’avant lors du cycle aquatique de leur bras gauche.
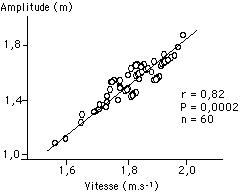
Figure : Relation linéaire entre la vitesse de nage et l’amplitude de déplacement de la hanche sur l’axe antéro-postérieur
Cette grande avancée vers l’avant du corps, et donc cette vitesse de nage importante peuvent-elles être expliquée par un certain type d’organisation spatio-temporelle des segments du membre supérieur ?
Nous avions déjà observé que les sujets observaient des durées totales, aquatiques et aériennes relativement semblables. Ce ne sont donc pas ces différents paramètres temporels qui peuvent expliquer la différence de vitesse de nage. Le tableau suivant met en évidence les relations entre les paramètres définis à partir des amplitudes et des durées obtenues par le poignet et le coude lors des différents points caractéristiques du cycle aquatique, et la vitesse de nage.
FIGURE : Coefficients linéaires des relations entre l’amplitude et la durée de différents paramètres du cycle aquatique du poignet et du coude et la vitesse de nage
|
Amplitude |
Durée |
|
|
S-E |
r = 0,41 P = 0,004 |
- |
|
A |
r = 0,53 P = 0,0001 |
- |
|
P |
r = 0,41 P = 0,004 |
- |
|
R |
r = -0,37 P = 0,01 |
- |
|
Rcoude |
r = -0,69 P = 0,001 |
- |
|
BE1 |
- |
r = -0,81 P = 0,0001 |
|
BI |
r = 0,63 P = 0,001 |
r = 0,61 P = 0,001 |
|
BE2 |
- |
- |
Globalement pour les axes antéro-postérieur et vertical, la vitesse de nage semble plus principalement liée aux paramètres spatiaux que temporaux, alors que sur l’axe transversal, la vitesse de nage est corrélée à la fois au spatial et au temporel.
Bien que significatifs, les coefficients de corrélation reflétant les relations entre les paramètres spatiaux sur l’axe antéro-postérieur (tels que l’avancée du poignet en début de cycle aquatique) et la vitesse de nage restent assez faibles. En observant la FIGURE suivante, caractéristique de ces relations, nous observons qu’une nouvelle notion d’amplitude optimale de déplacement pourrait se détacher. En effet, bien que les nageurs les moins performants soient toujours ceux qui enregistrent la plus faible amplitude de déplacement de leurs articulations, les meilleurs nageurs ne sont pas toujours représentés par les plus fortes amplitudes. A partir d’un certain niveau de performance, il y aurait un certain plafonnement au niveau des valeurs d’amplitude.

Figure : relation linéaire entre la vitesse de nage et les coordonnées maximales atteintes au cours de l’avancée du poignet en début de cycle aquatique
Cette augmentation générale de l’amplitude avec le niveau de performance peut être le fait direct d’un mouvement du poignet et du coude vers l’avant, mais également d’un mouvement d’extension de l’épaule vers l’avant et/ou d’une translation globale du corps. Afin d’observer précisément la conséquence sur la technique de nage de cette augmentation d’amplitude, les trajectoires du poignet et du coude ont été superposées à celle du corps lors d’un cycle aquatique (FIGURE).
A la vue du schéma précédent, l’augmentation de l’amplitude de mouvement serait l’effet direct d’une translation globale du corps vers l’avant. En effet en début de cycle aquatique, entre l’entrée de la main dans l’eau et le point d’avancée maximal sur l’axe antéro postérieur par exemple, le corps avance parallèlement au mouvement du poignet et du coude.
Sur l’axe transversal, la durée du premier balayage externe du poignet est lié négativement à la vitesse de nage. cela signifie que les meilleurs nageurs sont ceux qui placent leur articulation très rapidement dans l’axe du corps. A très haut-niveau, cette première phase deviendrait pratiquement inexistante du point de vue de sa durée, le poignet étant placé dès l’entrée dans l’eau.
La phase de balayage interne correspondant sur l’axe antéro-postérieur à la phase d’avancée vers l’avant, et sur l’axe vertical à la phase précédent l’atteinte de la profondeur maximale, est très lié à la vitesse de nage, que ce soit au niveau de son amplitude ou de sa durée. Les meilleurs nageurs sont ceux qui enregistrent la plus grande amplitude de mouvement sur une durée relativement élevée.
La dernière phase du cycle aquatique (phase de second balayage externe) n’est pas en relation avec la vitesse de nage. Ce résultat semble être en désaccord avec les études antérieures qui préconisaient souvent de pousser loin en arrière avec la main avant sa sortie de l’eau. Les résultats des meilleurs sprinters actuels ne semblent accorder d’importance à cette dernière phase.
Sur l’axe vertical, les meilleurs nageurs augmentent la profondeur de leurs appuis de la même manière qu’ils enregistrent de plus grandes amplitudes antéro-postérieures.
En résumé : notre étude tend à montrer que les meilleurs sprinters actuels sont caractérisés par :
- sur l’axe antéro-postérieur :
* une augmentation de l’avancée du poignet en début de cycle aquatique
* un faible recul du poignet et du coude
- sur l’axe transversal :
* une courte phase de prise d’appui
* une augmentation simultanée de l’amplitude et de la durée de la phase de balayage vers l’intérieur
- sur l’axe vertical :
* une augmentation de la profondeur des appuis, notamment au niveau du poignet.